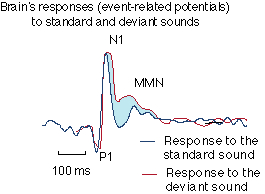
According to a review page on the Helsinki University Psychology Department's web site,
The mismatch negativity (MMN) is a component of the auditory event related potential (ERP) which is elicited task-independently by an infrequent change in a repetitive sound. The MMN can be recorded in response to any discriminable change in the stimulus stream.
Here is a picture illustrating the effect:
Note that the "negative" direction is upward in this plot, as is common for ERP displays. As always with ERP, these are averages of repeated recordings aligned at stimulus onset.
MMN was apparently first reported by Näätänen, Gaillard, and Mäntysalo (1978).
The corresponding phenomenon in MEG experiments is sometimes called MMNm, or sometimes "Mismatch Field" (MMF).
Of course, there is considerable intrinsic interest in the exact nature of the phenomenon, its neural implementation, etc., and in applications to problems from coma prognosis to audiometrics. Here I'm highlighting the reasons why it might be useful in studying the perception of conspecific vocalizations.
The effect is apparently independent of attention and of subject task, and works (sometimes) even in sleeping or sedated subjects, which in principle should make it easier to use with children and animals. The effect has been documented in various animals (monkeys, cats, guinea pigs), and in newborn human infants.
The effect is a fairly early one (100-200 ms, anyhow < 250 ms), and the generator is located in primary auditory cortex. Animal studies find MMN effects in the thalamus (medial geniculate nucleus). Despite this relative automaticity, it can be affected by some kinds of learning, both long-term (vowel categories in different languages) and short-term (training for unfamiliar VOT categories).
In humans, MMN/MMF is lateralized differently for speech than for non-speech stimuli. The details (though complex and full of loose ends) suggest early "modular" segregation of the sound stream -- in primary sensory cortex or even in subcortical (e.g. thalamic) circuits.
Animal studies claim to have isolated the cell populations and neurotransmitters involved.
The technique is relatively sensitive -- roughly, it seems that if two auditory stimuli can reliably be discriminated, then they will create the MMN/MMF effect.
ERP apparatus is inexpensive, somewhat portable and easily available (though MEG is not).
Some concerns:
Brief auditory stimuli are presented at regular intervals. Most (typically 85-90%) of the stimuli are of one type ("standards"), while a random selection of (10-15% of) the stimuli are of a different type ("deviants"). Stimuli are about 10 to 300 ms long. Interstimulus intervals tend to be .5 seconds to about 2.5 seconds. As the ISI lengthens, the effect decreases. Increasing rarity of the deviant stimuli produces stronger effects.
In some (but not all) versions of the paradigm, assignments of stimuli as standards or deviants is counterbalanced. When this is not done, there is obviously a potential confounding with SRP phenomena.
Subjects typically do not attend to the stimuli, but rather watch silent movies or read. In some cases, "ignore" blocks are matched with blocks in which the subjects count the deviants.
The differences between standards and deviants may be frequency, duration, intensity, timbre or some combination. Speech differences (isolated vowels or initial consonants in CV syllables) also work, as do interaural time differences, order differences between stimulus subparts, absence of an expected stimulus in the sequence, and so forth.
For MMN, several hundred presentations are typically averaged, whereas for MMF, several thousand are -- due to SNR issues in each case.
Subjects need not (and typically do not) attend to the stimuli. The effect has been demonstrated in subjects who are sleeping or sedated.
Apparently the one thing subjects should not be is relaxed and happy -- thus Surakka et al. 1998 found that "MMN was very similar during text reading and neutral and negative slide viewing, but was significantly attenuated during the viewing of positively valenced and little arousing visual emotional stimuli. We suggest that these stimuli served as a biological signal of a non-threatening and non-appetitive environment."
According to the previously cited MMN web page
MMN is elicited by both increments and decrements in basic stimulus features (for a thorough review, see Näätänen, 1992). The MMN, however, is not elicited when a stimulus sequence begins or, similarly, when stimuli are presented with very long interstimulus intervals (ISIs). Thus, it was concluded that no stimulus per se is an adequate stimulus for the MMN generator mechanism, as the system responds to the difference between the consecutive stimuli. This response pattern is clearly separable from the behavior of N1 response; the N1 amplitude is largest in response to the first stimulus of a series, strongly attenuating thereafter and showing only partial recovery to a subsequent different stimulus.
The observations that even decrements in stimulus intensity and, especially, in duration elicited the MMN need to be stressed because they provide strong evidence that the MMN is not a reflection of "fresh" neural activity in tonotopically and/or ampliotopically organized cortex (i.e., whereby the responses elicited by deviant stimuli would be generated in a location spatially separate from that of responses elicited by standard stimuli).
MMN apparently must be distinguished from the general effects of "oddball" presentation paradigms. Thus Goldstein and Aldrich (1999) write (p. 92):
Terms commonly used for two early (<250 ms) ERPs are processing negativity (PN) and mismatch negativity (MMN), proposed by Näätänen and his colleagues ... These terms have psychophysiologic connotations that make them difficult to compare across other ERPs (e.g. P300, which has a temporal dimension), and across AEPs in general.
and p. 95:
The P300 is so prominent [in oddball-paradigm recordings] that often all differences between the frequent and oddball AEPs are genericized by many as P300. Even the paradigm by means of which the ERPs are elicited often is called the P300 paradigm.
It appears that the P300 response is attention-dependent, however. (Goldstein and Aldrich p. 101) explain that the normal audiometric oddball procedure involves focusing subjects' attention on the oddballs
A common procedure is to have the subject ... count silently the number of oddball signals presented during a test trial. The listener is rewarded verbally or monetarily for the correctness of the count reported at the end of the trial. The subject obviously must be awake and attentive during the trial.
P300 and other late ERPs can be elicited from inattentive subjects who are not instructed to count the oddball. Resulting ERPs, however, are small or even indistinguishable in this ignore condition ...
ERPs occurring before 300 ms are less attention-dependent ...
The figure below (from Misulis 1994, p. 214) exemplifies the expected P300 pattern:
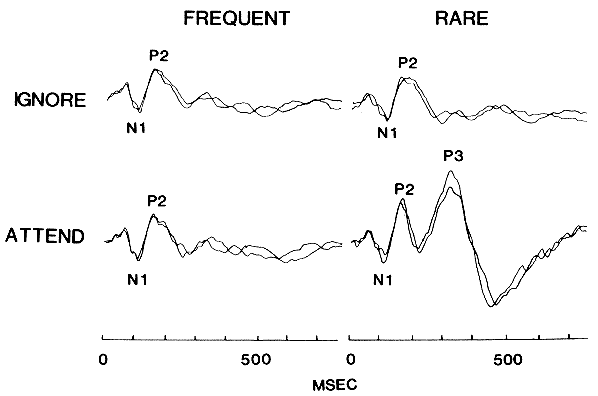
Here "FREQUENT' means "standard" and "RARE" means "deviant", in the terminology we've been using. "IGNORE" means a a block of trials in which the subjects are not paying attention to the deviants, and may be performing some distracting task (the source does not say). "ATTEND" means a block of trials in which the subject is counting deviant stimuli. In all cases, the plotted traces are averages of all trials for a given combination of stimulus type and subject task. I believe that the overlaid traces are different subjects. In this figure, negative polarity is plotted downwards on the y-axis. I presume that what is labelled "P3" here is what is referred to as "P300" in the quotes above.
Note that the MMN response (if it is present here) should exist in the region between N1 and P2, in the "IGNORE" condition. That is, the trace on the upper right should show MMN relative to the trace on the upper left. This mode of plotting the data makes it hard to see if an MMN effect is there or not (let's hope it's there somewhere!). The point is that the effects of conscious attention to the oddball stimuli seem to be very much larger than the MMN response, at least in examples like this one. Thus in an fMRI or PET "oddball" experiment, the analogue of the MNN response would have to be clearly different in spatial localization from P300 and other, since there is no chance to separate it out in time.
There are said to be different localizations of MMN or (especially) MMF generators depending on stimulus characteristics. However, I have not read many of these studies carefully, and remain unclear as to whether they adequately control for the possibility that stimulus differences are contributing in a way that is independent of their role in the mismatch detection process as such.
Here is the abstract of a summary article:
Cerebral generators of mismatch negativity (MMN) and its magnetic counterpart (MMNm) elicited by sound changes. Alho, K. Ear Hear, (1995) 16:1 38-51.
Infrequent (deviant) sounds occurring in a sequence of repetitive (standard) sounds elicit an event-related brain potential (ERP) response called the mismatch negativity (MMN) even in the absence of attention to these sounds. MMN appears to be caused by a neuronal mismatch between the deviant auditory input and a sensory-memory trace representing the standard stimuli. This automatic mismatch process has presumably a central role in discrimination of changes in the acoustic environment outside the focus of attention. Thus, localizing cerebral generators of MMN might help identify brain mechanisms of auditory sensory memory and involuntary attention. This review summarizes results from studies aimed at localizing MMN generators on the basis of (1) scalp-distribution, (2) magnetoencephalographic (MEG), (3) intracranial, and (4) brain-lesion data. These studies indicate that a major MMN source is located in the auditory cortex. However, the exact location of this MMN generator appears to depend on which feature of a sound is changed (e.g., frequency, intensity, or duration), as well as on the complexity of the sound (e.g., a simple tone versus complex sound). Consequently, memory traces for different acoustic features, as well as for sounds of different complexity, might be located in different regions of auditory cortex. However, MMN appears to have generators in other brain structures, too. There is some evidence for contribution of frontal-lobe activity to the MMN, which might be related to the involuntary switching of attention to a stimulus change occurring outside the focus of attention. In addition, intracranial MMN recordings in animals suggest that at least in some species, MMN subcomponents also may be generated in the thalamus and hippocampus.
A 1998 animal study (cats) by Karmos et al. reports that "the first surface positive ERP component displayed tonotopic distribution above the A I area, while the amplitude maximum of the MMN appeared above the A II area and had only minor frequency dependence". This study also concludes that "the MMN represent a local disinhibition in the supragranular layers of the A II area".
A 1998 human MEG study by Rinne et al. reports that a frontal component of MMN, "observed in about half the 15 subjects", "peaked at 5-20 ms later than the temporal component", consistent with the hypothesis that this component "is associated with involuntary switching of attention to stimulus change".
Several studies report that MMN/MMF for non-speech stimuli is greater in the right (nondominant) hemisphere. One study (Levanen, S, Ahonen, A, Hari, R, McEvoy, L, Sams, M. Cereb Cortex, (1996) 6:2 288-96) found bilateral MMF in the supratemporal cortex (i.e. primary auditory cortex), but found that "two temporally overlapping but spatially separate sources, one in the temporal lobe and another in the inferior parietal cortex, were necessary to explain the right-hemisphere MMF".
Several studies have found that MMN/MMF for speech sounds (in human subjects, natch) is greater in the left (dominant) hemisphere. One MEG study ("Noise-induced shift in hemispheric lateralization of speech processing", Yury Shtyrov, Teija Kujala, Jyrki Ahveninen, Mari Tervaniemi, Paavo Alku, Risto J. Ilmoniemi and Risto Näätänen; Neuroscience Letters, 251 141-144, 1998) found that for speech "in clear" (presumably about 50DB SNR?), there was essentially no right-hemisphere MMF response; as background white noise was introduced, the right-hemisphere MMF responses increased. At 10 DB SNR (not very bad, really), the right hemisphere MMF dominated.
An attempt to use dichotic presentation to selectively engage left- and right-hemisphere MMN responses (Praamstra, P, Stegeman, DF. Electroencephalogr Clin Neurophysiol, (1992) 82:1 67-80) failed, leading to the conclusion that "bilateral MMN generators [are] symmetrically involved" in the usual (nonspeech) case.
In a series of studies using synthetic vowel stimuli, the MMN response was enhanced when the deviant stimulus was a "prototype" vs. when it was not. Specifically, in Language-specific phoneme representations revealed by electric and magnetic brain responses. Naatanen, R, Lehtokoski, A, Lennes, M, Cheour, M, Huotilainen, M, Iivonen, A, Vainio, M, Alku, P, Ilmoniemi, RJ, Luuk, A, Allik, J, Sinkkonen, J, Alho, K. Nature, (1997) 385:6615 432-4, the standard stimulus was always a version of /e/ allegedly equally good for Finnish and Estonian, whereas the deviant stimulus was either one of a number of other Finnish vowel prototypes, or a vowel /õ/ that occurs in Estonian but not in Finnish. Finnish subjects showed reduced MMN response to the Estonian vowel, whereas Estonians did not.
On the other hand, [Acoustic versus phonetic representation of speech as reflected by the mismatch negativity event-related potential. Sharma, A, Kraus, N, McGee, T, Carrell, T, Nicol, T. Electroencephalogr Clin Neurophysiol, (1993) 88:1 64-71] found that pairs of stimuli taken from a /da/-/ga/ continuum were equally good at eliciting NNM regardless of whether they were within-category or cross-category. [Note: this is a bit hard to square with the general observation that the more discriminable the stimuli, the greater the MMN effect; it's hard to believe that the within-category stimuli were are discriminable as the across-category stimuli -- unless they were also a lot closer in acoustic terms].
Several studies have found correlations between individual differences in discrimination performance (measured psychophysically) and MMN magnitudes (citations TK).
A study by Lyytinen et al. found group differences in MMN responses to stimuli varying in vowel and consonant lenght between infants "born to familially dyslexic families" and a control group.
A study by Ceponiene et al. found group differences in MMN amplitude in 7-9-year-olds, for groups split on the basis of pseudo-word repetition and pseudo-word learning.
There are said to be controls in some studies that show that MMN is not just a consequence of habituation causing a lessened response (somewhere in auditory cortex) to the standard stimuli. For instance, the responses to deviant stimuli are said to differ crucially from the responses to block-initial stimuli. I haven't seen the details of these studies. However, MMN boosters and users clearly believe that the efect represents the result of a comparison of an acoustic memory trace (of one or more previous "standard" stimuli) against the current "deviant" stimulus.
I have not seen a fully (or even mostly) explicit model about just what is being compared to what, however.
The closest thing to an explicit model seems to be that proposed by Ritter and Winkler, and I haven't yet read their paper(s).
Detection of stimulus deviance within primate primary auditory cortex: intracortical mechanisms of mismatch negativity (MMN) generation. Javitt, DC, Steinschneider, M, Schroeder, CE, Vaughan, HG, Jr, Arezzo, JC. Brain Res, (1994) 667:2 192-200.
Mismatch negativity (MMN) is a cognitive, auditory event-related potential (AEP) that reflects preattentive detection of stimulus deviance and indexes the operation of the auditory sensory ('echoic') memory system. MMN is elicited most commonly in an auditory oddball paradigm in which a sequence of repetitive standard stimuli is interrupted infrequently and unexpectedly by a physically deviant 'oddball' stimulus. Electro- and magnetoencephalographic dipole mapping studies have localized the generators of MMN to supratemporal auditory cortex in the vicinity of Heschl's gyrus, but have not determined the degree to which MMN reflects activation within primary auditory cortex (AI) itself. The present study, using moveable multichannel electrodes inserted acutely into superior temporal plane, demonstrates a significant contribution of AI to scalp-recorded MMN in the monkey, as reflected by greater response of AI to loud or soft clicks presented as deviants than to the same stimuli presented as repetitive standards. The MMN-like activity was localized primarily to supragranular laminae within AI. Thus, standard and deviant stimuli elicited similar degrees of initial, thalamocortical excitation. In contrast, responses within supragranular cortex were significantly larger to deviant stimuli than to standards. No MMN-like activity was detected in a limited number to passes that penetrated anterior and medial to AI. AI plays a well established role in the decoding of the acoustic properties of individual stimuli. The present study demonstrates that primary auditory cortex also plays an important role in processing the relationships between stimuli, and thus participates in cognitive, as well as purely sensory, processing of auditory information.
Demonstration of mismatch negativity in the monkey. Javitt, DC, Schroeder, CE, Steinschneider, M, Arezzo, JC, Vaughan, HG, Jr. Electroencephalogr Clin Neurophysiol, (1992) 83:1 87-90.
In humans, deviant auditory stimuli elicit an event-related potential (ERP) component, termed mismatch negativity (MMN), that reflects the operation of a cortical detector of infrequent stimulus change. Epidural auditory ERPs were recorded from 3 cynomolgous monkeys in response to soft and loud clicks. Oddball loud or soft stimuli elicited a long-duration frontocentral negativity, peaking at approximately 85 msec, that was superimposed upon cortically generated obligatory ERP components. These data suggest that monkeys might serve as a heuristically valuable system in which to study the neurochemical and neuroanatomical substrates of early context-dependent ERP generation.
Role of cortical N-methyl-D-aspartate receptors in auditory sensory memory and mismatch negativity generation: implications for schizophrenia. Javitt, DC, Steinschneider, M, Schroeder, CE, Arezzo, JC. Proc Natl Acad Sci U S A, (1996) 93:21 11962-7.
Working memory refers to the ability of the brain to store and manipulate information over brief time periods, ranging from seconds to minutes. As opposed to long-term memory, which is critically dependent upon hippocampal processing, critical substrates for working memory are distributed in a modality-specific fashion throughout cortex. N-methyl-D-aspartate (NMDA) receptors play a crucial role in the initiation of long-term memory. Neurochemical mechanisms underlying the transient memory storage required for working memory, however, remain obscure. Auditory sensory memory, which refers to the ability of the brain to retain transient representations of the physical features (e.g., pitch) of simple auditory stimuli for periods of up to approximately 30 sec, represents one of the simplest components of the brain working memory system. Functioning of the auditory sensory memory system is indexed by the generation of a well-defined event-related potential, termed mismatch negativity (MMN). MMN can thus be used as an objective index of auditory sensory memory functioning and a probe for investigating underlying neurochemical mechanisms. Monkeys generate cortical activity in response to deviant stimuli that closely resembles human MMN. This study uses a combination of intracortical recording and pharmacological micromanipulations in awake monkeys to demonstrate that both competitive and noncompetitive NMDA antagonists block the generation of MMN without affecting prior obligatory activity in primary auditory cortex. These findings suggest that, on a neurophysiological level, MMN represents selective current flow through open, unblocked NMDA channels. Furthermore, they suggest a crucial role of cortical NMDA receptors in the assessment of stimulus familiarity/unfamiliarity, which is a key process underlying working memory performance.
Nonprimary auditory thalamic representation of acoustic change. Kraus, N, McGee, T, Littman, T, Nicol, T, King, C. J Neurophysiol, (1994) 72:3 1270-7.
1. The mismatch response, or mismatch negativity (MMN), is a neurophysiologic response to stimulus change. In humans and other animals, the MMN may underlie the ability to discriminate acoustic differences, a fundamental aspect of auditory perception. 2. This study investigated the role of the thalamus in the generation of a tone-evoked MMN in guinea pigs. Electrodes were placed in the caudomedial (nonprimary) and ventral (primary) subdivisions of the auditory thalamus (medial geniculate nucleus). Surface epidural electrodes were placed at the midline and over the temporal lobe. The MMN was elicited by a deviant stimulus (2,450-Hz tone burst) embedded in a sequence of standard stimuli (2,300-Hz tone bursts). 3. A tone-evoked MMN was present in nonprimary thalamus but was absent in the primary thalamus. Surface-recorded MMNs were measured at the midline but not over the temporal lobe. The correspondence between nonprimary thalamic responses and midline surface potentials, and between primary thalamic responses and temporal surface potentials, is consistent with data reported for the auditory middle latency responses in guinea pigs. 4. The results demonstrate that the nonprimary auditory thalamus contributes to the generation of a tone-evoked MMN in the guinea pig. Furthermore, the data indicate that the guinea pig is a feasible model for investigating central auditory processes underlying acoustic discrimination.
Evoked potential correlates of stimulus deviance during wakefulness and sleep in cat--animal model of mismatch negativity. Csepe, V, Karmos, G, Molnar, M. Electroencephalogr Clin Neurophysiol, (1987) 66:6 571-8.
Auditory evoked potentials (EPs) elicited by standard (STs) and deviant tones (DTs) of different probabilities were studied in freely moving cats during wakefulness and sleep. A large double peaked negativity, so-called mismatch negativity (MMN), was evoked by the unattended low probability DTs. The EPs recorded from the AI and AII areas of the auditory cortex showed more dynamic changes than the vertex and association cortical responses. The amplitude of the MMN was inversely proportional to the probability of DTs. The latency of the MMN showed dependence both on the location of the recording site and on the probability of DTs. During slow wave sleep (SWS) the MMN of increased latency could be evoked only at the lowest probabilities. The cortical distribution of the MMN changed in the SWS.